引言
初乳是一种特殊的物质。母牛初乳(MC)的成分与普通牛奶截然不同;事实上,它们的差异如此显著,以至于我们将产后24小时内乳腺分泌的液体称为“非牛奶”物质。 初乳含有大量免疫球蛋白(Ig),尤其是IgG,这些物质源自母牛妊娠最后几周的血液循环。此外,初乳还含有许多其他蛋白质,包括激素、酶和生长因子,这些成分可能影响新生犊牛的新陈代谢。同时,初乳中大量脂肪、维生素和矿物质能满足犊牛出生后立即的营养需求。
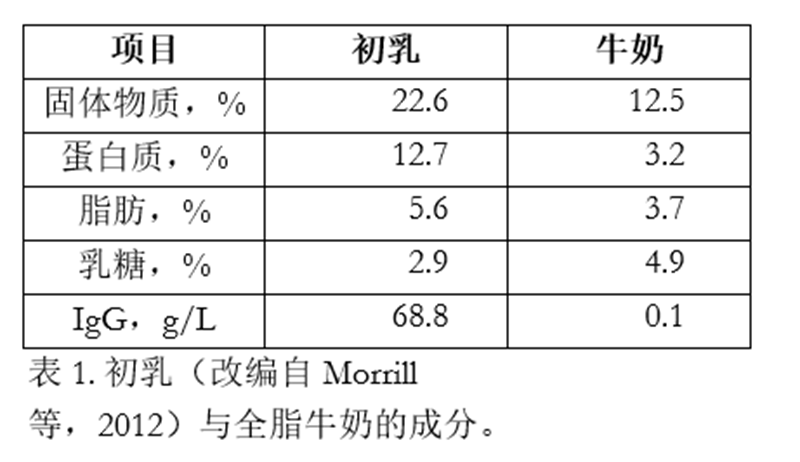
表1对比了初乳与全脂牛奶的成分。显而易见,初乳比牛奶更浓稠,且固体物质和总营养素含量远高于牛奶。
那么,当犊牛摄入一餐初乳时会发生什么?假设犊牛在出生后第一小时内被喂食推荐剂量的一加仑(3.8升)初乳。我们为犊牛提供了多少营养?这些营养又会如何被利用?表2将一加仑(3.8升)初乳与一加仑全脂牛奶进行了对比。
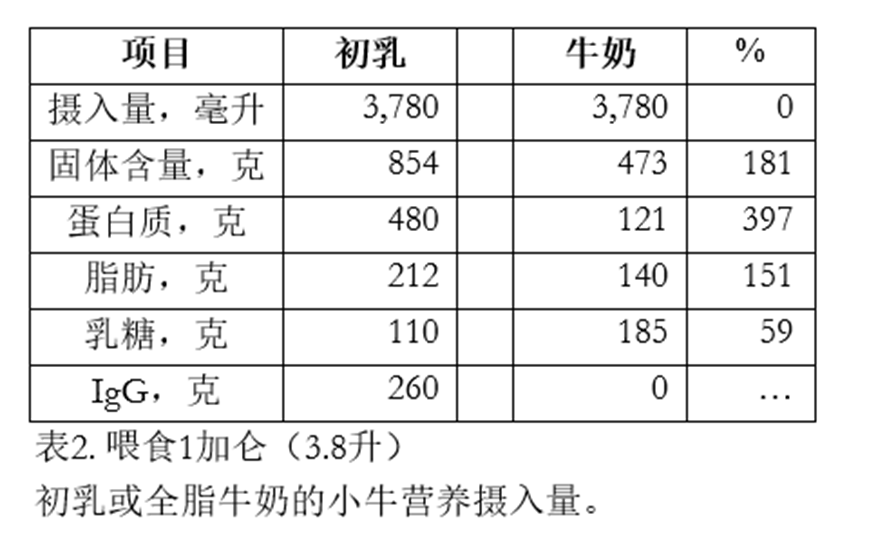
一头犊牛摄入3.8升初乳(蛋白质含量为12.7%),将摄入总计480克蛋白质。相比之下,若犊牛饮用同等量的全脂牛奶,蛋白质摄入量仅为121克。一顿大份的初乳提供的蛋白质约为同等份量牛奶的4倍!
当然,我们的目标是向新生犊牛提供IgG,使其免受环境病原体的侵害。但其余的蛋白质会发生什么呢?这是一个有趣的观察结果,或许能为新生犊牛的新陈代谢提供一些见解。
我们可以通过计算一项名为“IgG吸收表观效率”(AEA)的统计指标,来监测新生犊牛对IgG的吸收情况。该指标反映了喂食初乳中的IgG在犊牛出生24小时后被血液吸收的效率。 要计算这一指标,我们需要知道犊牛的体重(用于估算血容量)、出生24小时后的血浆(或血清)IgG浓度,以及犊牛摄入的IgG总量。计算公式如下:
AEA (%) = 血浆中IgG浓度 (g/L) × 血浆容量 (L) ÷ IgG摄入量 (g)
通常我们使用体重(BW)来估算血浆容积:
AEA (%) = 血浆中IgG浓度 (g/L) × [体重 (kg) × 9%] ÷ IgG摄入量 (g)
因此,一头血浆IgG浓度为12 g/L、体重40 kg且摄入150克IgG的小牛,其IgG吸收效率为:
AEA (%) = 12 × [40 × 0.09] ÷ 150 = 0.288,即 28.8%
通常,IgG的AEA范围在20%至约35%之间(Quigley和Drewry,1998)。
如果IgG的吸收效率为28%,那么其他蛋白质的情况如何?我们能否计算出非IgG蛋白质的AEA?答案是肯定的,但首先我们需要一些额外信息。
犊牛出生时血液中没有循环IgG,但其血液中含有非IgG蛋白。新生犊牛出生后,血液中的蛋白质含量约为4.0 g/dl(40 g/L)。这些蛋白质包括转运蛋白、白蛋白以及许多其他蛋白质。然而,其中不含(或仅含极少量)IgG。
要推算非IgG蛋白质的变化情况,我们需要估算出生时血液中的总蛋白含量——不仅要考虑浓度(即4.0 g/dl),还要考虑蛋白质的总克数。我们可以根据血浆容量进行估算,但遗憾的是,血浆容量是动态变化的。 也就是说,出生24小时后的血浆量(根据已发表的研究,我们假设其约为体重的9%,参见Quigley等,1998)与出生时的血浆量并不相同。当犊牛饮用初乳后,血浆量会增加。因此,我们假设出生时血浆量略低于9%。 有人估计出生时的血浆量更接近体重的7%。
如果我们在出生时采集了血液样本以测定总蛋白,我们还可以测定血细胞比容或红细胞压积。我们假设,尽管血浆量会随着初乳的摄入而增加,但循环系统中细胞的总数变化甚微(注:这一假设几乎可以肯定是不正确的,但就我们进行的计算而言,误差程度应该很小)。
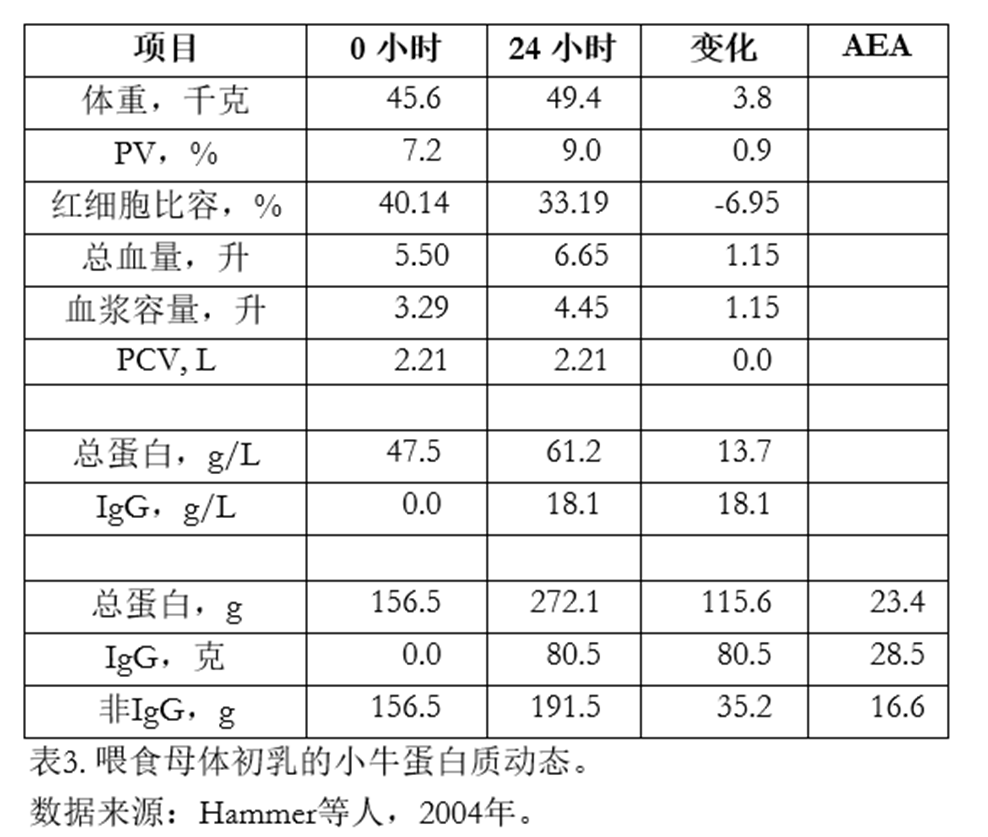
请看表3。这些数据来自一组每天喂食3.8升MC的小牛(Hammer等人,2004)。 出生 24 小时时,犊牛体重为 49.4 公斤,红细胞比容 (PCV) 为 33.19%,血清总 蛋白和 IgG 浓度分别为 6.15 g/dl 和 18.1 g/L。 我们假设犊牛在出生24小时时的血浆容积(PV)为9%。因此,其总血浆容积(以升为单位)为4.45 L。利用红细胞比容(PCV)数据,我们估算红细胞容积为2.21 L。
假设从出生0小时到24小时,红细胞总量未发生变化(出生0小时和24小时均为2.21 L),则可计算出该犊牛的血浆量和总血容量。 该犊牛出生时血清总蛋白浓度为4.75 g/dl(47.5 g/L;表3),24小时时为6.12 g/dl(61.2 g/L)。总蛋白的净变化量为13.7 g/L,这归因于IgG及其他蛋白质的吸收。
由此可计算出犊牛出生时及24小时后血液中的蛋白质含量(单位:克)。从表3可见,犊牛出生时血液中含有156.5克蛋白质;24小时后增至272.1克。即增加了115.6克。 犊牛从初乳中摄入了494克总蛋白质,因此115.6 / 494 = 23.4%的利用率。 也就是说,在犊牛出生24小时后,其血液中约有23%的摄入蛋白质得以保留。
我们对IgG(24小时血浆中80.5克 / 摄入量282克 = 28.5%的利用率)和非IgG蛋白(通过差值计算:35.2克 / 212克 = 16.6%的利用率)也进行了同样的计算。
值得注意的是,非IgG部分的吸收效率较低——相比于吸收效率约为29%的IgG,非IgG蛋白部分的吸收效率仅约为17%。那么,其余的蛋白质都去哪儿了呢?
尽管我们假设许多初乳蛋白能逃过肠道消化,但有研究表明,在最初的24小时内,肠道中至少会发生部分消化。 Yvon等人(1993)报告称,新生羔羊体内α-乳清蛋白的大部分蛋白质会在瘤胃和小肠中被消化。因此,肠道中确实存在部分蛋白质的消化过程,这意味着某些蛋白质被分解为单个氨基酸,这些氨基酸可能用于蛋白质合成或葡萄糖生成。然而,免疫球蛋白对消化具有更强的抵抗力。
非IgG蛋白(及部分IgG)也可能被肌肉和肝脏代谢以产生葡萄糖,这些葡萄糖随后可用于体温调节及其他功能。其他循环中的 蛋白可能被机体用于蛋白质合成,从而离开血液循环。最后,部分蛋白质会被肾脏滤过并随尿液排出。这种被称为蛋白尿的状况在幼龄犊牛中很常见。
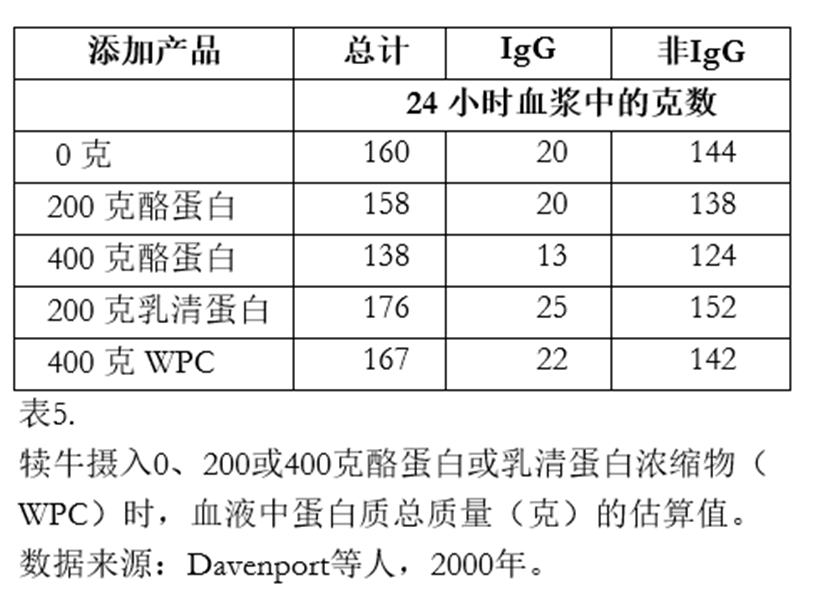
似乎随着摄入的非IgG蛋白量增加,其利用效率反而降低。Davenport等人(2000)给犊牛喂食了添加了0、200或400克酪蛋白或乳清蛋白的初乳补充剂。
表4显示,随着非IgG蛋白质摄入量的增加,非IgG组分的AEA值呈下降趋势。喂食添加了400克酪蛋白或乳清蛋白粉的小牛,其非IgG组分的AEA值分别为2.1%和7.8%。这表明小牛可能存在蛋白质摄入的上限。 表5显示了出生24小时后血浆中蛋白质的估计质量(包括总蛋白、IgG和非IgG蛋白)。看来,额外喂给犊牛的蛋白质已被代谢并用作能量,或者可能被机体排出了。 Davenport等人(2000)也报告称,随着蛋白质的添加,尿素氮浓度有所增加,这表明他们喂食的非IgG蛋白中至少有一部分被机体代谢了。
总结
初次喂食初乳的犊牛,特别是喂食3.8升时,会摄入大量蛋白质——这可能是其一生中任何其他时期摄入量都无法比拟的。其中一部分蛋白质会在胃肠道内被消化为氨基酸和小肽。部分蛋白质将以完整形式被吸收;这些蛋白质可能会被代谢为氨基酸,或者如果血液中蛋白质浓度过高,则可能随尿液排出。
参考文献
Davenport, D. F., J. D. Quigley, III, J. E. Martin, J. A. Holt, and J. D. Arthington. 2000. 向初乳或初乳补充剂中添加酪蛋白或乳清蛋白对新生犊牛IgG吸收的影响。 《乳业科学杂志》 83:2813–2819.
Hammer, C. J., J. D. Quigley, L. Ribeiro, and H. D. Tyler. 2004. 含IgG浓缩物和生长因子的初乳替代品及初乳补充剂的特性分析。《乳业科学杂志》87:106–111.
莫里尔,K. M. 2012. 改进现有实验室方法以快速测定初乳IgG浓度及新生犊牛对初乳IgG的吸收。《博士学位论文》,爱荷华州立大学,艾姆斯。
奎格利,J. D.,与 J. J. 德鲁里。1998。产前及产后母牛向犊牛的营养与免疫物质转移。《乳业科学杂志》81:2779–2790。
Quigley, J. D., J. J. Drewry, 和 K. R. Martin. 1998. 荷斯坦牛和泽西牛犊血浆容积的估算。《乳业科学杂志》81:1308–1312. 伊冯,M.,D. 勒维厄,M. C. 瓦吕,J. P. 佩利西耶和P. P. 米兰德。1993. 新生羔羊初乳蛋白质的消化。《营养学杂志》123:586-596.