Clic aquí para una versión en PDF
Introducción
La nutrición y el manejo de los terneros parecen dividirse en dos períodos distintos: el primer día de vida y todo lo posterior. Todos sabemos la importancia del Día 1, el período más importante en el que los terneros recién nacidos pueden absorber inmunoglobulinas en el torrente sanguíneo sin digestión. Este es el período en el que la alimentación con calostro (o el uso de suplementos/reemplazos de calostro) es esencial para proporcionar al ternero inmunidad pasiva y nutrientes críticos para la supervivencia.
Pero, después de las primeras 24 horas de vida, la mayoría de las lecherías y ranchos de terneros trasladan a los recién nacidos, que ahora tienen entre 24 y 36 horas de edad, al alojamiento y manejo “normal” de la operación de crianza de terneros. Por supuesto, estos “bebés” pueden recibir un trato especial: enseñarles a beber de la tetina o del balde, vacunas, etc. Pero, en su mayor parte, asumimos que este joven animal puede ser alimentado y manejado como todos los demás terneros.
Entonces, aquí está la pregunta: ¿el ternero de un día es capaz de digerir, absorber y utilizar nutrientes como sus hermanos y hermanas de varias semanas de edad? ¿Es su sistema inmunológico tan competente para combatir patógenos como el de terneros mucho mayores? La respuesta corta es no y no. El objetivo de esta Nota sobre terneros es explicar algunas de las diferencias y proponer que podríamos considerar un enfoque diferente para el manejo de terneros del “Día 2”. Consideraremos diferencias en tres áreas: flora intestinal (bacterias), digestión y metabolismo e inmunidad.
Flora bacteriana. Cuando el ternero nace por primera vez, el tracto digestivo es estéril. Durante las primeras 24 horas de vida, aproximadamente, las bacterias se instalan en el tracto, colonizando desde “ambos extremos” (boca y ano) y, finalmente, se pueden encontrar grandes cantidades de bacterias en todo el tracto.
A las 24 horas de vida, hay una población considerable en el tracto, pero muchas de estas bacterias son “transitorias”, es decir, se establecen en el intestino porque fueron inoculadas a través de los alimentos o del medio ambiente. Normalmente no se encuentran (al menos en concentraciones mensurables) en el intestino de los terneros mayores. En las décadas de 1940 y 1950 se trabajó mucho para comprender los cambios en las poblaciones bacterianas en el rumen de los terneros recién nacidos. Es probable que se produzcan cambios similares en el intestino de los terneros jóvenes; sin embargo, hay menos datos disponibles.
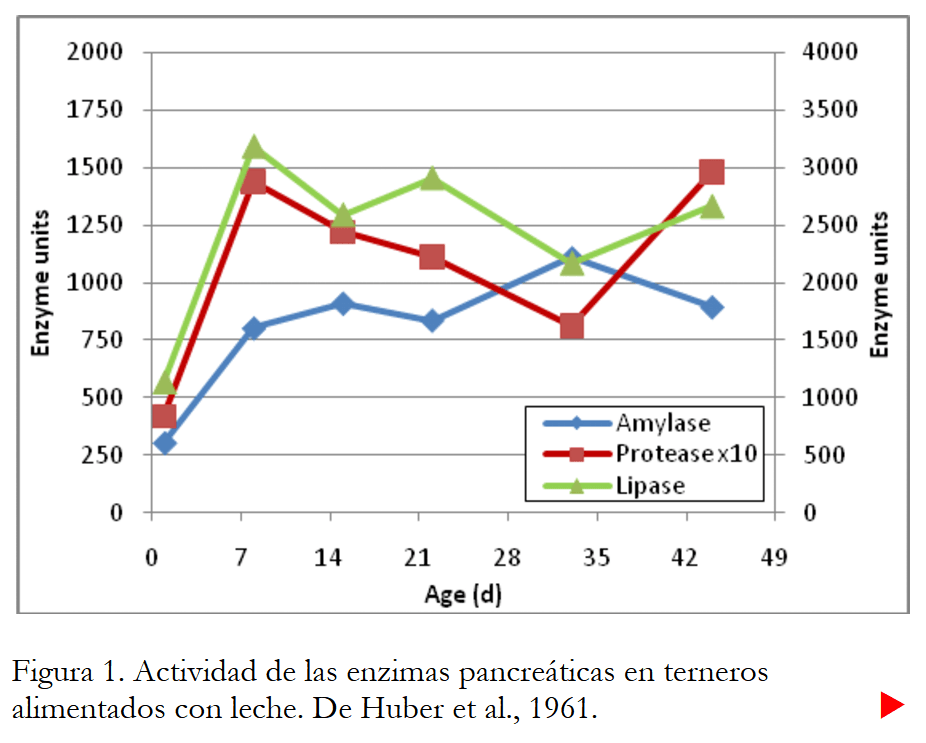
Digestión y metabolismo. Las enzimas y secreciones gastrointestinales que utilizan los terneros para digerir su alimento no se activan “mágicamente” a las 24 horas de edad. Muchos o la mayoría de ellos aumentan gradualmente con el tiempo. Por ejemplo, Huber et al. (1961) midieron la actividad enzimática en terneros alimentados con leche de 1 a 44 días de edad. La actividad media de las enzimas en el páncreas se muestra en la Figura 1. Como puede ver, la actividad de todas las enzimas (lipasa, proteasa y amilasa) fue menor el día 1 que otros días y luego aumentó el día 8. A partir de entonces, las actividades no cambiar dramáticamente. Otros investigadores han informado datos similares (Ternouth y Buttle, 1973; Ternouth et al., 1976; Sissons, 1981). Generalmente, las actividades de muchas (pero no de todas) las enzimas tienden a aumentar con el tiempo. Sin embargo, esto a menudo depende del tipo de dieta, la cantidad de alimento sólido consumido (y el posterior desarrollo del rumen) y la edad. Pero los datos parecen claros: la mayoría de las enzimas son menos activas en los primeros días de vida. Por lo tanto, no debemos asumir que los terneros están listos para digerir el mismo tipo de dieta el día dos que el día 20.
Inmunidad. La absorción de inmunoglobulinas se completa a las 24 horas de edad. El proceso de maduración intestinal (llamado cierre intestinal) que termina al final del primer día. Muchos otros componentes del sistema inmunológico todavía están deprimidos en el día 2: los niveles de complemento disminuyen en los primeros días. Otros aspectos de la respuesta inmune del ternero están menos desarrollados y requieren tiempo para madurar por completo. Rossi et al. (1981) y muchos otros han documentado la inmadurez del sistema inmunológico neonatal.
Un enfoque dietético
E
| Gravedad especifica | 1.056 | 1.035 | 1.033 | 1.032 |
| Solidos, % | 23.9 | 14.1 | 13.6 | 12.9 |
| grasa, % | 6.7 | 3.9 | 4.3 | 4.0 |
| SNF, % | 16.7 | 9.8 | 9.5 | 8.8 |
| Proteina, % | 14.0 | 5.1 | 4.1 | 3.1 |
| Lactosa, % | 2.7 | 4.4 | 4.7 | 5.0 |
| Ca, % | 0.26 | 0.15 | 0.15 | 0.13 |
| Caseina, % | 4.8 | 3.8 | 2.9 | 2.5 |
| Ig, g/L | 60 | 24 | N/A* | 0.09 |
| IgG, g/L | 32 | 15 | N/A* | 0.06 |
Tabla 1. Composición de la leche de vacas después del parto. Adaptado de Foley y Otterby, 1978. *N/A = no disponible. ntonces, si un ternero en el día 2 todavía no está “normalmente desarrollado” (al menos en comparación con un ternero de 14 a 21 días de edad), ¿cómo manejamos a los terneros para que se adapten a las diferencias en su metabolismo? ¿Son apropiados los métodos actuales de alimentación y manejo para los terneros del día 2? La respuesta corta es que no lo sabemos con certeza. Sin embargo, podemos observar cómo la naturaleza aborda el día 2 y tratar de aprender cómo nuestra nutrición y manejo podrían adaptarse para mejorar el rendimiento de los terneros.
La Tabla 1 contiene el contenido de nutrientes e inmunoglobulinas (Ig total e IgG) del calostro (primer ordeño después del parto) y la leche de transición recolectada los días 2 y 3. Estos se comparan con la leche normal (adaptado de Foley y Otterby, 1978).
Como podemos ver, la composición de la leche de transición varía significativamente tanto del calostro como de la leche madura en términos de sólidos, nutrición y contenido de inmunoglobulinas. contenido de IgG sigue siendo significativo. Según Foley y Otterby, el contenido de IgG es aproximadamente la mitad que el del calostro del primer ordeño. Si calculamos la ingesta de nutrientes e IgG (supongamos que un ternero consume 4 litros de calostro, leche de transición y leche madura), vemos que todavía hay muchas diferencias entre la ingesta de leche de transición del día 2, leche madura y calostro. Estas cifras se encuentran en la Tabla 2. En comparación con el calostro, la leche del día 2 contiene menos sólidos (-41%), grasa (-42%), proteínas (-74%), caseína (-21%), Ig total (-60% ) e IgG (-52%). Sin embargo, en comparación con el calostro, los terneros alimentados con leche del día 2 consumirán más lactosa (+63%) debido a los cambios en el contenido de lactosa de la leche de transición del día 2 en comparación con el calostro.
| Solidos, % | 956 | 564 | 544 | 516 |
| grasa | 268 | 156 | 172 | 160 |
| Lactosa | 108 | 176 | 188 | 200 |
| Proteína | 560 | 204 | 164 | 124 |
| Caseína | 192 | 152 | 116 | 100 |
| Ig | 240 | 96 | N/A* | <1 |
| IgG | 124 | 60 | N/A | <1 |
Cuadro 2. Ingesta de nutrientes, Ig e IgG en terneros alimentados con 4 litros de calostro, leche de transición y leche madura. Adaptado de Foley y Otterby, 1978. *No disponible.
Cuando comparamos la ingesta del día 2 con la ingesta de 4 litros de leche madura, vemos que los terneros alimentados con leche del día 2 comen más sólidos (+9%), proteínas (+65%), caseína (+52%), Ig e IgG. La ingesta de grasa es similar y la lactosa es ligeramente menor que cuando los terneros beben leche madura.
¿Cuáles son las implicaciones para esta información? Parece que los terneros en el día 2 consumirán más sólidos, proteínas e IgG cuando beben leche de transición en comparación con la leche entera. Blättler et al. (2001) y Bühler et al. (1998) informaron que alimentar con más calostro tenía efectos positivos sobre la maduración del intestino, la actividad enzimática y la digestión.
De particular interés es la alimentación continua de cantidades relativamente grandes de IgG. El día 2, los terneros alimentados con leche de transición todavía reciben más de 50 gramos de IgG. Si asumimos que las vacas lecheras proporcionan IgG en su calostro y leche por alguna razón, podríamos concluir que la alimentación continua con IgG confiere un beneficio continuo a los terneros incluso aunque no se absorban en el torrente sanguíneo. Y, de hecho, muchos artículos publicados sugieren que la alimentación continua de IgG y otras proteínas funcionales proviene del calostro. Por eso, muchas publicaciones de extensión recomiendan la alimentación continua con calostro durante los tres días posteriores al nacimiento:
- http://ag.udel.edu/anfs/faculty/kung/articles/importance_of_colostrum_for_calv.htm
- http://www.uky.edu/Ag/AnimalSciences/dairy/extension/nut00109.pdf
- http://www.farmllc.org/custom3.html
- http://www.rennut.com/articles/pdf/Newborn%20Calf%20Care.pdf
Desafortunadamente, muchos productores no tienen leche de transición disponible. Algunos venden su leche de transición; otros trasladan a los terneros de la lechería a un rancho de terneros el día 1 y es posible que solo tengan acceso a un sustituto de leche comercial.
Una posible solución es agregar una pequeña cantidad de calostro del primer ordeño o sustituto de calostro comercial al sustituto de leche.
Referencias
Anderson, K. L., T. G. Nagaraja, J. L. Morrill, T. B. Avery, S. J. Galitzer, and J. E. Boyer. 1987. Ruminal microbial development in conventionally or early-weaned calves. J. Anim. Sci. 64:1215-1226.
Blättler, U., H. M. Hammon, C. Morel, C. Philipona, A. Rauprich, V. Romé, I. Le Huërou-Luron, P. Guilloteau and J. W. Blum. 2001. Feeding colostrum, its composition and feeding duration variably modify proliferation and morphology of the intestine and digestive enzyme activities of neonatal calves. J. Nutr. 131:1256–1263.
Bühler, C., H. Hammon, G. L. Rossi, and J.W. Blum. 1998. Small Intestinal morphology in eight-day-old calves fed colostrum for different durations or only milk replacer and treated with long-R3-insulin-like growth factor I and growth hormone. J. Anim. Sci. 1998. 76:758–765.
Bryant, M. P., N. Small, C. Bouma, and I. Robinson. 1958. Studies on the composition of the ruminal flora and fauna of young calves. J. Dairy Sci. 41:1747-1766.
Foley, J. A., and D. E. Otterby. 1978. Availability, storage, treatment, composition, and feeding value of surplus colostrum: A review. J. Dairy Sci. 61:1033-1060.
Huber, J. T., N. L. Jacobson, R. S. Allen, and P. A. Hartman. 1961. Digestive enzyme activities in the young calf. J. Dairy Sci. 44::1494-1501.
Rossi C.R., G. K. Kiesel, R. S. Hudson, T. A. Powe, and L. F. Fisher. 1981. Evidence for suppression or incomplete maturation of cell-mediated immunity in neonatal calves as determined by delayed-type hypersensitivity responses. Am. J. Vet. Res. 42:1369-70.
Sissons, J. W. 1981. Digestive enzymes of cattle. J. Sci. Food Agric. 32:105-114.
Ternouth, J. H., and H. L. Buttle. 1973. Concurrent studies on the flow of digesta in the duodenum and of exocrine pancreatic secretion of calves. Br. J. Nutr. 29:387-397.
Ternouth, J. H., J.H.B. Roy, and S. M. Shotton. 1976. Concurrent studies on the flow of digesta in the duodenum and of exocrine pancreatic secretion of calves. 4. The effect of age. Br. J. Nutr. 36:523-535.