Introduction
The use of immunoglobulins (Ig) to reduce the effects of pathogenic challenge has been recognized for hundreds of years. In this situation, we are not talking about feeding antibodies to calves during the first 24 hours of life, but after the first 24 hours. Although these antibodies are not absorbed, they can help the immune system of calves in maintaining gastrointestinal health. We have all heard of the recommendation to feed calves colostrum for the first three days of life. The reason for feeding during days two and three are not because the antibodies will be absorbed into the bloodstream (they won’t), but because they “coat the gut” with antibodies that can help keep the calf healthy. NOTE: for additional information, see Calf Note #60.
To understand the role of Ig in replacing antibiotics, it is important to understand that the intestinal tract is the largest immunological organ in the body. The total area of these mucosal surfaces, which cover these tube-like tissues are at least two hundred times larger than those of skin (Takahashi and Kiyono, 1999). The large amount of lymph tissue (primarily as Peyer’s patches) in the gut also contributes to the immunological capability of the intestine. These tissues appear to be particularly important in enteric disease caused by viruses and bacteria (Frost et al., 1997; Brodersen and Kelling, 1999). Therefore, in addition to providing critical digestive functions, the intestine must also prevent diseases from entering the body.
The gastrointestinal tract is constantly exposed to insults consumed by the animal. These may include pathogenic organisms, toxins, noxious chemicals, physical insults (e.g., hardware disease) and many others. Organs in the gastrointestinal tract have many methods to deal with these insults, including secretion of digestive enzymes and acid, harboring of commensal organisms, and other methods (Kruzel et al., 1998). Of particular interest, however, is the presence of Ig in the intestines. The second component involves functional immunological elements found in the mucosal and submucosal compartments, e.g., gut associated lymphoid tissue. When gut integrity is disrupted by invasive pathogens or by trauma, a myriad of pro-inflammatory mediators are released from cells in the gut wall that exert actions in the tissue or gut lumen. Immunoglobulin is an important defense mechanism in overall immune response in the intestinal and production of Ig by gut associated lymphoid tissue is a critical function of these tissues.
Traditionally, the only Ig considered important in the intestine was IgA, which is produced by epithelial cells. Indeed, researchers continue to focus on production of intestinal IgA as a means of controlling disease (Sagodira et al., 1999; Coffin et al., 1999). However, other recent evidence suggests that IgG may also play an important role in reducing the risk of disease in animals. The two primary sources of IgG in the gut is through secretion of IgG from the blood into the intestine and oral consumption of IgG from milk or colostrum (lacteal secretions), blood or eggs.
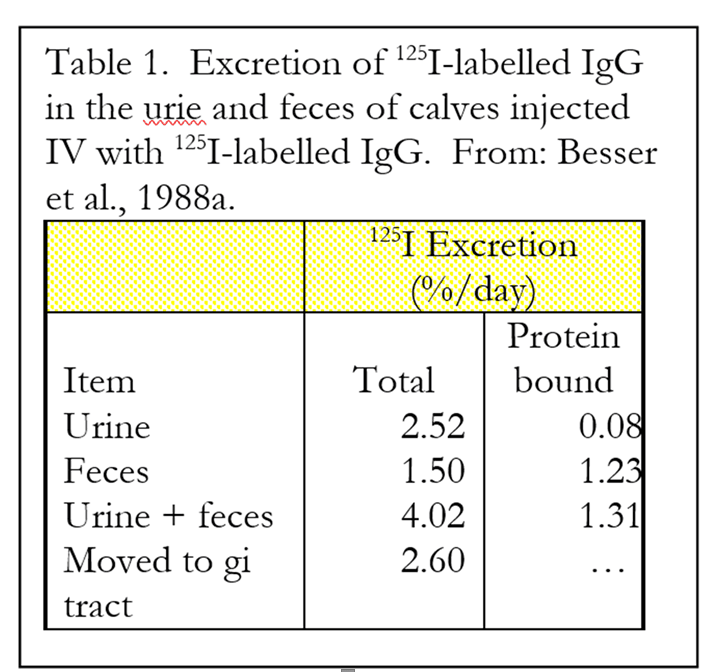
Movement of circulating IgG into the gut. Research done at Washington State University by Dr. Tom Besser and coworkers investigated the movement of circulating IgG into the intestinal tract and the role of IgG in reducing effects of microbial challenge (1988a, b). The researchers conducted two studies to determine the metabolic fate of IgG that entered the bloodstream. In the first study, calves were injected with a radioactive (125I) labeled IgG directly into the blood. The calves (n = 24) were colostrum deprived and obtained from a commercial U.S. dairy. The excretion of the radioactive label was then monitored over time by collecting urine and fecal samples and determining the amount of radiation they contained. The excretion of total radiation and the total radiation still bound to protein (an estimate of the “intact” IgG) were measured.
An average of 2.52% of the 125I was excreted in the urine every day (Table 1). Most of this was not bound to protein (only about 3% of urinary excretion), indicating that the IgG excreted in urine had been previously catabolized. Also, 1.5% of injected 125I was excreted by way of the feces. Most of this (82%) was still bound to protein, indicating that these IgG were not degraded prior to excretion in the feces. The total excretion of 125I was 4.02% per day of the amount injected. Regression analysis indicated that the half-life of the injected 125I containing IgG was 17.9 days.
Calves were euthanized and the amount of 125I was determined in various compartments of the intestine to estimate directly the amount of IgG that moved from the circulation into the intestine. The total values corresponded to a daily transfer of 2.60% of the total infused 125I into the gastrointestinal tract. Most of this IgG appears to be secreted into the intestine as intact IgG, but a portion apparently is degraded by intestinal enzymes. The authors estimated that if a calf were to consume and absorb 100 g of IgG from maternal colostrum within the first 24 hours, it would subsequently secrete 1 to 4 grams of IgG back into the intestine daily for the first two weeks of life.
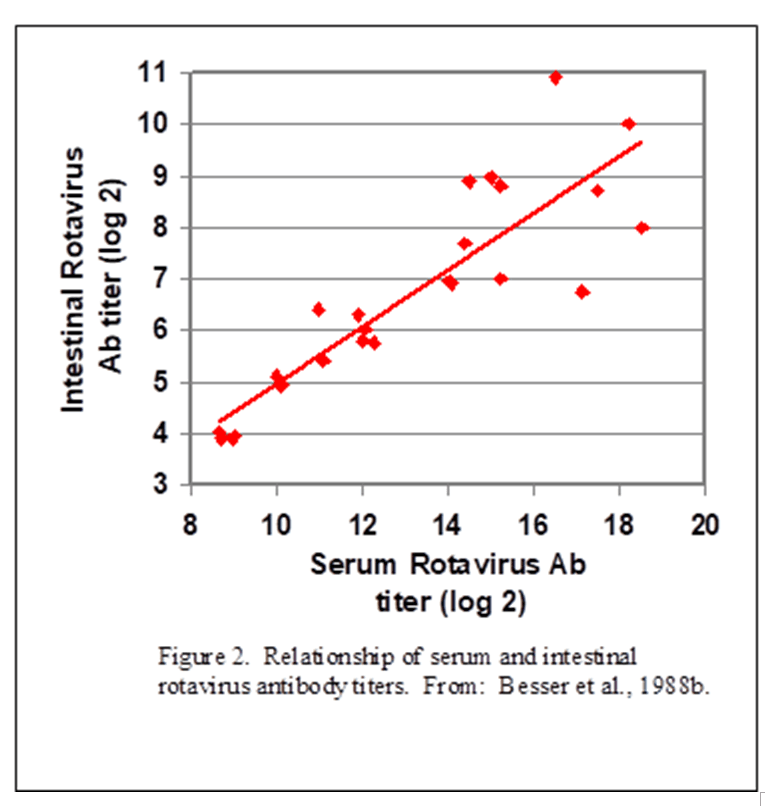
In a second experiment, Besser and coworkers (Besser et al., 1988b) fed newborn calves colostrum containing antibodies against a specific strain of rotavirus. Dry cows were immunized with a vaccine against the rotavirus at 6 and 3 weeks prior to expected calving to produce colostrum containing the specific antibody. The amounts of specific antibody were then measured in the blood and gastrointestinal contents following sacrifice at 5 or 10 days of age.
The correlation between serum rotavirus antibody and intestinal rotavirus antibody (Figure 1) showed a close correlation. This means that calves 1) absorbed the specific antibody from the colostrum consumed within the first 24 hours, 2) the specific antibodies then moved from the circulation into the lumen of the intestine, and 3) the movement of specific antibodies into the intestine occurred in proportion to concentrations in the blood.
The value of intestinal IgG. Many bacteria and viruses that infect calves are enteric – typically causing intestinal damage and signs of disease (diarrhea, dehydration). Immunoglobulins in the intestine could assist the animal to mount an effective immune response when they attach to the antigenic binding sites on the specific pathogen. Therefore, movement of IgG from the circulation into the intestinal lumen would be one way to provide immunity in response to the pathogens that infect the animal by the fecal-oral route.
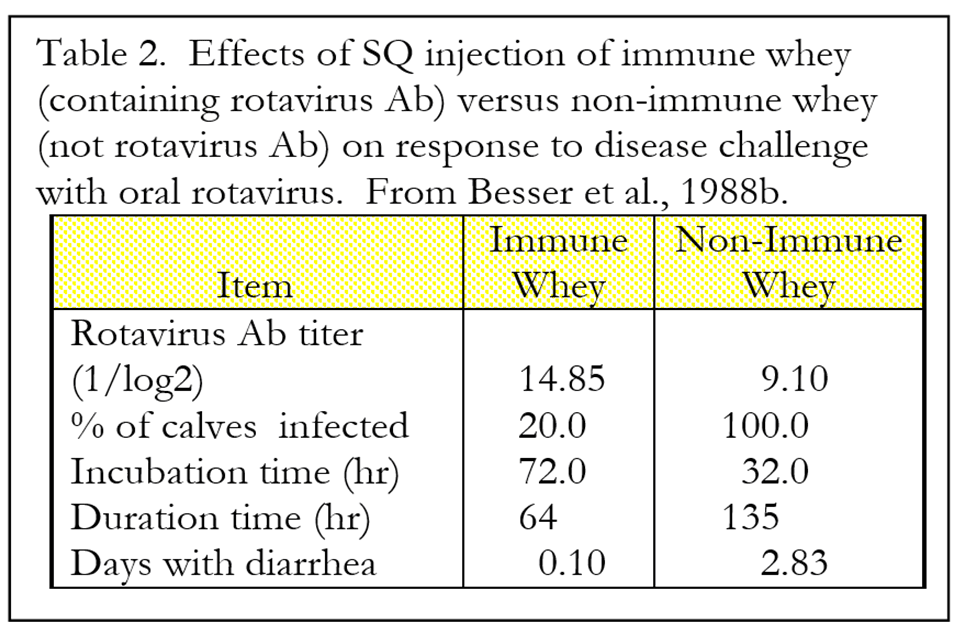
To determine if there is any value to circulating IgG in dealing with intestinal pathogens, Besser and coworkers injected calves subcutaneously with 1.25 liters of whey extracted from the colostrum of cows immunized against rotavirus or colostrum from non-immunized cows. The control group was fed colostrum from non-immunized cows. These calves were then challenged with enteropathogenic strain of rotavirus at 72 and 96 hours after birth.
Administration of IgG by subcutaneous injection protected calves against rotavirus infection (Table 2). Calves treated with subcutaneous “immune” whey (whey containing rotavirus antibody) had higher serum antibody titers against rotavirus and were more protected against oral rotavirus challenge than calves that were injected with “non-immune” whey. Presumably, the mode of action for the immune whey was via movement of the IgG from the circulation into the intestinal lumen, where the rotavirus was present. It is important to note that these calves were fed no colostrum, so the only source of antibody was through subcutaneous injection.
These studies indicate that:
- Ig in the intestine play an active role in the resistance to pathogenic organisms that infect calves via the oral route, such as rotavirus.
- Ig in the intestine are sufficiently resistant to digestion to provide immune response. Studies have documented the relative resistance of IgG to proteolytic degradation in the gut.
- A major source of IgG in the intestine of newborn calves is from circulating IgG that are absorbed from ingestion of colostrum within the first 24 hours.
- Larger concentrations of IgG in the serum generally produce larger concentrations of IgG in the lumen of the intestine.
Reduced digestibility of Ig. Immunoglobulins are more resistant to proteolysis than many other proteins. This is necessary for IgG to provide local response in the intestine of the animal. Roos et al. (1995) reported that the recoveries of N of ingested IgG and IgM still immunologically active were 19±3% and 19±4%, respectively in human patients consuming 15N labeled preparations of Ig. According to the data of Roos et al., the ileal digestibility of IgG in healthy humans was 79%. Interestingly, much of the immunological activity was associated with the F(ab’)2 fragments, which are produced by pepsin and trypsin activity on IgG. The F(ab’)2 fragments contain a molecular weight of ~100 kDa.
Researchers measured serum levels of rotavirus specific maternally derived antibodies in neonatal pigs. Pigs were grouped into non-detectable, low, or high serum titers. Pigs were then challenged with virulent rotavirus at 3 days of age and monitored for infection and disease. All inoculated pigs shed rotavirus and developed diarrhea, and pigs with highest levels of circulating antibody to rotavirus developed less severe diarrhea and shed rotavirus for fewer days than pigs with lower antibody titers. The researchers concluded that circulating maternal antibody plays a significant role in mitigating clinical disease and movement of antibodies from the circulation into the lumen of the intestine is important in this response. Apparently, the movement of antibodies from the circulation into the intestine is important to animal health.
Future Calf Notes will look at the three primary sources of antibodies, milk and colostrum, blood and eggs. Each has advantages and disadvantages and research with each source of antibodies has provided insight into the value of antibodies in calf health.
References
- Besser, T. E., T. C. McGuire, C. C. Gay, and L. C. Pritchett. 1988a. Transfer of functional immunoglobulin G (IgG) antibody into the gastrointestinal tract accounts for IgG clearance in calves. J. Virology. 62:2234-2237.
- Besser, T. E., C. C. Gay, T. C. McGuire, and J. F. Evermann. 1988b. Passive immunity to rotavirus infection associated with transfer of serum antibody into the intestinal lumen. J. Virology. 62:2238-2242.
- Brodersen, B.W. and C. L. Kelling. 1999. Alteration of leukocyte populations in calves concurrently infected with bovine respiratory syncytial virus and bovine viral diarrhea virus. Viral Immunol. 12:323-334.
- Coffin S. E., S. L. Clark, N. A. Bos, J. O. Brubaker, and P. A. Offit. 1999. Migration of antigen-presenting B cells from peripheral to mucosal lymphoid tissues may induce intestinal antigen-specific IgA following parenteral immunization. J. Immunol. 163:3064-3070.
- Frost, A. J., A. P. Bland and T. S. Wallis. 1997. The early dynamic response of the calf ileal epithelium to Salmonella typhimurium. Vet. Pathol. 34:369-386.
- Kruzel M. L., Y. Harari, C. Y. Chen, and G. A. Castro. 1998. The gut. A key metabolic organ protected by lactoferrin during experimental systemic inflammation in mice. Adv. Exp. Med. Biol. 443:167-173.
- Roos, N., S. Mahe, R. Benamouzig, H. Sick, J. Rautureau, and D. Tome. 1995. 15N-labelled immunoglobulins from bovine colostrum are partially resistant to digestion in human intestine. J. Nutr. 125:1238-1244.
- Sagodira S., S. Iochmann, M. N. Mevelec, I. Dimier-Poisson, and D. Bout. 1999. Nasal immunization of mice with Cryptosporidium parvum DNA induces systemic and intestinal immune responses. Parasite Immunol.199921:507-516.
- Takahashi, I. And H. Kiyono. 1999. Gut as the largest immunologic tissue. J. Parenter. Enteral Nutr. 23(Suppl.):S7-12.